Kap. 11B. Opprinnelsen til livet: Vitenskapsfolk kaster terning
(oversatt fra 'Should Christians embrace evolution' (2009), kap. 11 av John C. Walton -forskningsprofessor em. i kjemi ved St. Andrews, Skotland)
Teorier for selvorganisering
 Vitenskapsfolk har vært klar over ekstremt lav sannsynlighet for at liv skal oppstå ved tilfeldighet i årtier, men håper enda på en naturalistisk vei ut av uføret. Wikipedia nevner over 15 scenarier, hvorav noen er overlappende, som er foreslått for livets opprinnelse. Og denne listen er ikke utfyllende. (15) Eksistensen av så mange konkurrerende teorier er i seg selv bevis for at de alle er beheftet med seriøse problemer. Tidlig i debatten ble det foreslått at aminosyrer eller kanskje nukleotider, kan ha iboende fysio-kjemiske egenskaper som disponerte dem til å selv-organisere på biologisk nyttige måter. Interessen i denne sammenheng fikk et stort oppsving, da Dean Kenyon og Gary Steinman oppdaget betydelig kjemiske preferanser i assosiering mellom aminosyrer. I deres bok 'Biolkjemis predestinasjon' foreslo de at disse ulikhetene i affinitet førte til at noen sekvenser ble foretrukket framfor andre, og at denne selektiviteten i sin tur kan ha ledet til de samme sekvensene som ble funnet i fungerende proteiner. (16)
Vitenskapsfolk har vært klar over ekstremt lav sannsynlighet for at liv skal oppstå ved tilfeldighet i årtier, men håper enda på en naturalistisk vei ut av uføret. Wikipedia nevner over 15 scenarier, hvorav noen er overlappende, som er foreslått for livets opprinnelse. Og denne listen er ikke utfyllende. (15) Eksistensen av så mange konkurrerende teorier er i seg selv bevis for at de alle er beheftet med seriøse problemer. Tidlig i debatten ble det foreslått at aminosyrer eller kanskje nukleotider, kan ha iboende fysio-kjemiske egenskaper som disponerte dem til å selv-organisere på biologisk nyttige måter. Interessen i denne sammenheng fikk et stort oppsving, da Dean Kenyon og Gary Steinman oppdaget betydelig kjemiske preferanser i assosiering mellom aminosyrer. I deres bok 'Biolkjemis predestinasjon' foreslo de at disse ulikhetene i affinitet førte til at noen sekvenser ble foretrukket framfor andre, og at denne selektiviteten i sin tur kan ha ledet til de samme sekvensene som ble funnet i fungerende proteiner. (16)
Mens teorien vokste i popularitet, begynte Kenyon selv å tvile på den og forkastet den til slutt. Oppfølgeren til denne type selvorganisering, ville være at kjemisk pre-determinerte aminosyrer og sekvenser, på en eller annen måte ble overført til nukleotide-sekvenser i DNA. Imidlertid viste eksperimentelle studier at affiniteten påvist av Kenyon og Steinman ikke korrelerte med aktuelle sekvens-mønstre i kjente proteiner. (17) Det var heller slik at funksjonelle proteinsekvenser hadde blitt dannet av informasjon kodet i DNA, og ikke var en konsekvens av noen kjemisk styrt samling av aminosyrer. Informasjonsflyten var FRA DNA TIL proteiner og ikke motsatt.
Bilde 1. Hydrogenbinding mellom baser
Det fantes enda en mulighet for at selv-organisering og informasjonsdannelse kunne finne sted fra egenskaper i nukleotidene. Alle fire nukleotider inneholder samme type sukker og fosfat-enheter. De skiller seg bare i sin sammesetning av baser. Når de lenker seg sammen for å forme nukleinsyre, så knytter deres sukker og fosfatenheter seg sammen og hver sammenknytning er samme type kjemiske bånd, som knytter sammen suksessive sukkermolekyler i en polynukleotide. (BILDE) Det er ingen kontakt melllom base-sammensetningene for tilgrensende nukleotider. Fra et kjemisk synspunkt syntes det derfor alltid usannsynlig at det kunne være noen spesiell affinitet, som predisponerte nukleotider til å gruppere seg i spesielle sekvenser. Heller ikke har noen selektiv sammensetning av nukleotider noensinne vært observert. Biokjemikeren Bernd-Olaf Küppers konkluderte: "Egenskapene ved nukleinsyre indikerer at alle mulige kombinasjoner, fra et kjemisk synspunkt, er ekvivalente." (18)
 Michael Polany var den første som viste vei til en klar innsikt i gåten om informasjon knyttet til DNA. (19) Han tiltrakk oppmerksomhet til forskjellen mellom orden og informasjon. Om bokstaver i et alfabet alltid ble tvunget til å opptre i visse sekvenser, så kunne de bare overføre ett begrenset sett meninger. Han argumenterte at DNA's evne til å overføre informasjon faktisk avhang av den ikke-bundne evne nukleotidene har til å koble seg sammen i hvilken sekvens som helst, fri for kjemisk eller fysisk påvirkning. Om sammensetnings egenskapene til nukleotidene favoriserte visse sekvenser, så ville DNA's evne til å befordre informasjon bli alvorlig skadelidende. Han konkluderte: "Samme hva som er kilden til sammensetningen av DNA, så kan det bare fungere som kode fordi rekkefølgen deres ikke er resultat av potensiell energi. Det må være like fysisk ubestemt som rekkefølgen av ord på en skrevet side." Således er det ingenting ved egenskapene til verken aminosyrene eller nukleotidene som predisponerer dem til å samle seg i bestemte uttrykk, som start på selvorganisering. Ideen om at liv er forutbestemt eller uungåelig, får ingen støtte overhodet fra studiet av sammensetning av proteiner eller nukleinsyre. Den biokjemiske predestinasjonsteorien er nå genelt avskrevet.
Michael Polany var den første som viste vei til en klar innsikt i gåten om informasjon knyttet til DNA. (19) Han tiltrakk oppmerksomhet til forskjellen mellom orden og informasjon. Om bokstaver i et alfabet alltid ble tvunget til å opptre i visse sekvenser, så kunne de bare overføre ett begrenset sett meninger. Han argumenterte at DNA's evne til å overføre informasjon faktisk avhang av den ikke-bundne evne nukleotidene har til å koble seg sammen i hvilken sekvens som helst, fri for kjemisk eller fysisk påvirkning. Om sammensetnings egenskapene til nukleotidene favoriserte visse sekvenser, så ville DNA's evne til å befordre informasjon bli alvorlig skadelidende. Han konkluderte: "Samme hva som er kilden til sammensetningen av DNA, så kan det bare fungere som kode fordi rekkefølgen deres ikke er resultat av potensiell energi. Det må være like fysisk ubestemt som rekkefølgen av ord på en skrevet side." Således er det ingenting ved egenskapene til verken aminosyrene eller nukleotidene som predisponerer dem til å samle seg i bestemte uttrykk, som start på selvorganisering. Ideen om at liv er forutbestemt eller uungåelig, får ingen støtte overhodet fra studiet av sammensetning av proteiner eller nukleinsyre. Den biokjemiske predestinasjonsteorien er nå genelt avskrevet.
Bilde 2. Binær kode i tankepartikler
 RNA-verden
RNA-verden
Ved midten av 1980-tallet var det oppdaget at visse RNA molekyler, kalt ribosymer, kunne oppføre seg lik ensymer og katalysere noen viktige celle-reaksjoner. Grunnet dens store likhet til DNA, kunne RNA potensielt lagre genomisk informasjon ved hjelp av sine nukleotide-sekvenser, siden RNA også er sammensatt av lineære sekvenser av nukleotider. Ideen grep fatt i forestillingen om en prebiotisk verden, der RNA eller muligens pre-RNA (22) kunne påta seg begge roller til informasjonslagring så vel som spesifikk katalyse. Håpet var at en dermed kunne unngå kylling-egg problemet som mellom proteiner og DNA. I RNA-verdenens hypotese, oppsto ett RNA-molekyl som var i stand til å kopiere seg selv tilfeldig, ved kombinasjon av ribose, fosfat og sammensatte baser i en ursuppe. Kompleksiteten og funksjonaliteten økte gradvis, som følge av naturlig seleksjon. I neste stadium begynte RNA-molekylet å lage proteiner, og endelig dukket DNA opp, oppnådde den genetiske informasjonen og overtok som lagringsmedium, på grunn av deres større stabilitet. Dette scenariet er for tiden vel ansett i det vitenskapelige samfunn og sett som den mest lovende veien framover for forskning om livets opprinnelse.
Bilde 3. RNAs byggestener.
Det er imidlertid mange svakheter og usannsynligheter i teorien, som har hindret den i å oppnå universell anerkjennelse. Det er ingen kjente kjemiske måter for abiogenisk dannelse av nukleotider som trengs for RNA-oppbygging. De sammensatte basene er ustablie ved aktuelle temperaturer for scenarier for livets opprinnelse (23). Reaksjonen som antas å ha dannet ribose-sukker, produserer ikke sukker i nærvær av nukleinsyrer eller aminosyrer (24). Ribose er bare en av mange slags sukker dannet i reaksjonen, og ribose inneholder selv fire reaktive områder. Seleksjon av korrekte sukker-fosfat-lenker, under prebiotiske forhold, ville være et enormt problem. Alternative strategier for RNA-dannelse, utenom behov for fritt sukker, har vært utforsket i laboratorier. I disse eksperimentene ble nukleotidene dannet via syntetiske sekvenser. Men det problematiske opphavet til disse under prebiotiske forhold, og behovet for mangfoldige renselses-steg, gjør dette til en usannsynlig prebiotisk vei til RNA-nukletotider.
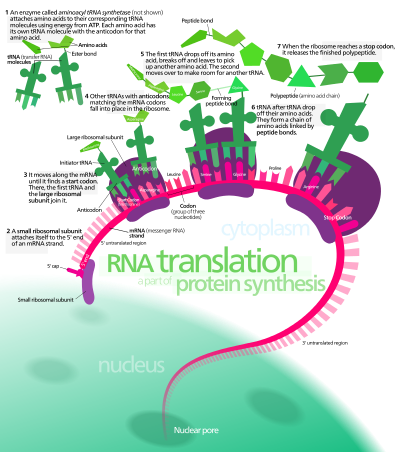 RNA molekyler er i seg selv mindre stabile enn DNA-molekyler og blir lett utsatt for hydrolyse. Skrøpeligheten til store RNA-molekyler gjør at de lett mister sin konsistens. Å lagre store informasjonsmengder i RNA for lengre tidsrom, er ikke et gjennomførbart forslag.RNA-hypoteser går ut fra spesifikke, operasjonelle RNS-sekvenser, som allerede er fungerende. Dermed forblir opprinnelsen til disse uløst. Det første selvreplikerende RNA-molekyl, måtte ha formet seg ved tilfeldig, ikke-styrt kjemiske kombinasjoner av nukleotider. Det antas at dette har skjedd, selv om oddsene er veldig høye for at det er slik. Videre er RNA-molekylene forestilt å starte å danne proteiner. Men hypotesen gir ingen forklaring på hvor informasjonen for dette, kommer fra. RNA-hypotesen har stimulert mye interesse omkring RNA-molekyler. Man søker å produsere ribozymer med bedre katalyse-muligheter og i stand til å utføre bedre stoffskifte. I noen forsøk forsøker en å endre sekvensene av naturlig forekommende RNA-katalysatorer. I andre screenes RNA-molekyler for spesifikke typer av katalyserende aktivitet. Så screenes resultatet, og de mest aktive plukkes ut. Prosessen gjentas inntil optimal katalyse-aktivitet er oppnådd. Det er tydelig at en oppnår seleksjon under styring av forskerteamet. Uten tvil har resultater som er av interesse fra et biokjemisk ståsted, blitt oppnådd ved å smugle inn seleksjon. Som vi tidligere har sett, må det eksistere en selvreplikerende organisme om det skal finne sted i naturen. Men ved hjelp av dette, har en dannet RNA-molekyler, som kan kopiere små seksjoner av seg selv. Det har vært produsert RNA-polymerase som kan forlenge eksisterende sekvenser med opptil 20 nukleotider (25). Likedan med korte RNAs som kan katalysere dannelse av peptid-bånd mellom amionsyrer (26). Dette har vært hyllet i populær-pressen som tegn på at kunstige livsformer snart vil dannes i prøverør (27).
RNA molekyler er i seg selv mindre stabile enn DNA-molekyler og blir lett utsatt for hydrolyse. Skrøpeligheten til store RNA-molekyler gjør at de lett mister sin konsistens. Å lagre store informasjonsmengder i RNA for lengre tidsrom, er ikke et gjennomførbart forslag.RNA-hypoteser går ut fra spesifikke, operasjonelle RNS-sekvenser, som allerede er fungerende. Dermed forblir opprinnelsen til disse uløst. Det første selvreplikerende RNA-molekyl, måtte ha formet seg ved tilfeldig, ikke-styrt kjemiske kombinasjoner av nukleotider. Det antas at dette har skjedd, selv om oddsene er veldig høye for at det er slik. Videre er RNA-molekylene forestilt å starte å danne proteiner. Men hypotesen gir ingen forklaring på hvor informasjonen for dette, kommer fra. RNA-hypotesen har stimulert mye interesse omkring RNA-molekyler. Man søker å produsere ribozymer med bedre katalyse-muligheter og i stand til å utføre bedre stoffskifte. I noen forsøk forsøker en å endre sekvensene av naturlig forekommende RNA-katalysatorer. I andre screenes RNA-molekyler for spesifikke typer av katalyserende aktivitet. Så screenes resultatet, og de mest aktive plukkes ut. Prosessen gjentas inntil optimal katalyse-aktivitet er oppnådd. Det er tydelig at en oppnår seleksjon under styring av forskerteamet. Uten tvil har resultater som er av interesse fra et biokjemisk ståsted, blitt oppnådd ved å smugle inn seleksjon. Som vi tidligere har sett, må det eksistere en selvreplikerende organisme om det skal finne sted i naturen. Men ved hjelp av dette, har en dannet RNA-molekyler, som kan kopiere små seksjoner av seg selv. Det har vært produsert RNA-polymerase som kan forlenge eksisterende sekvenser med opptil 20 nukleotider (25). Likedan med korte RNAs som kan katalysere dannelse av peptid-bånd mellom amionsyrer (26). Dette har vært hyllet i populær-pressen som tegn på at kunstige livsformer snart vil dannes i prøverør (27).
Bilde 4. Mange forutsetninger må gjøres for at proteiner kan dannes kun via RNA
Selv om ribosom-engineering har oppnådd bemerkelsesverdige resultater, så er det et faktum at den store majoritet av stoffskifte oppgaver ennå krever aminosyre-baserte enzymer, og ribosomer som er i stand til å erstatte dem, er ikke oppdaget. Videre er det flere alvorlige mangler ved påstanden om at ribosom-engineering er en gyldig modell for prebiotisk produksjon av selvreplikerende organismer. For det første spilte forskerteamet en kritisk rolle, da intelligente moekylær-biologer designet og gjennomførte eksperimentene. For det andre ble betydelige mengder av informasjonen som kreves for fungerende RNA, ble forsynt av intelligent seleksjon av opprinnelige RNA-sekvenser og i kontroll av eksperiment-betingelsene. For det tredje, så innebærer metoden med gjentatte screenings-faser at naturlig seleksjon er smuglet inn bakveien i eksperimentet. Ribosom-eksperimentene gir ikke forklaring på et naturlig opphav til livet. Tvert i mot viser resultatene at gitt et stort mål av input fra intelligente forskere og tilvirkede ingeniør-prosesser, og målstyring av screenings-prosesser, så kan framgang oppnås i å katalysere aktive sekvenser og replikasjon formes. I det input fra menneskelig intelligens har økt, så har balansen til bevisene vippet i favør av teorien at reproduksjon og liv oppsto fra intelligent design.
Evolusjonære algoritmer
Richard Dawkins har prøvd å la det naturlige utvalg tre inn til erstatning for algoritmebegrepet. Men det naturlig utvalg er ikke-algoritmisk og er følgelig en avsporing i stedet for en erstatning. Dawkins prøver basere sine 'beviser' bl.a. på et dataspill hvor mener å kunne vise hvordan orden blir til av kaos ved mutasjoner og naturlig utvalg. Han starter med den meningsløse bokstav-kombinasjonen:
- WDLM NTL DTJBK WIRZREZLMQ CO P, som han mener å omskape til høyverdig litteratur i løpet av 41 'generasjoner'. I hver generasjon gjør han en tilfeldig endring av en bokstav, og tester hvor nær han er kommet den setningen han skal til: METHINKS IT IS LIKE A WEASEL. Ut fra resultatet enten beholder eller forkaster han den siste versjonen. Det kjennetegner jo naturlig utvalg at en beholder det beste, når en skal mellom den gamle og den nye setningen. MEN Dawkins smugler inn all informasjonen selv, ved å sette opp Hamlet  setningen som en målskive, og avgjøre framskritt i forhold til avvik i forhold til den. I stedet for å illustrere effekten ved naturlig utvalg, gjør han følgende:
setningen som en målskive, og avgjøre framskritt i forhold til avvik i forhold til den. I stedet for å illustrere effekten ved naturlig utvalg, gjør han følgende:
i) Han illustrerer at liv må ha vært forhåndsprogrammert og brakt inn i systemet
ii) Han illustrerer at naturlig utvalg alltid snylter på eksisterende orden
iii) Han illustrerer at mutasjoner ikke skaper informasjon, Mutasjoner kan ikke bringe oss nærmere et mål med mindre dette målet er gitt på forhånd.
iv) Han illustrerer at biologisk orden ikke følger av død natur
v) Han illustrerer at livet ikke kan være resultat av en tilfeldig prosess
Oppsummert om Dawkins: han prøvde vise at livet har tilfeldig opphav og tilfeldig målsetting. Han oppnådde å vise at livet har ikke-tilfeldig opphav og ikke-tilfeldig målsetting. Det pussige er at berømmelse ble bygd på et bokprosjekt som grunnstøter så kraftig i ett av de første kapitlene. Men ofte er darwinister ikke i stand til å se at de jukser, for det finnes få korrektiv i deres omgivelser.
Kilder:
15. Abiogenese: https://en.wikipedia.org/wiki/Abiogenesis
16. D. Kenyon og G.Steinman, Biochemical Predestination (New York: McGraw-Hill, 1969)
17. R.A.Kok, J.A.Tayloer et al, 'A Statistical Examination of Self-Ordering', Origins of Life and Evolution of Biospheres 18 (1988), ss.135-142.
18. Bernd-Olaf Küppers, 'On the Prior Probability of the Existence of Life', in L. Krüger et al, The Probababalistic Revolution, vol. 2 (Cambridge, MA: MIT Press, 1987), ss.355-369.
19. M. Polanyi, 'Life's Irreducible Structure', Science 160 (1968), s.1308.
20. Se Meyer, Signature in the Cell, kap.14, s296 for en grundig diskusjon av RNA-verden
22. PNA (Peptide nukleinsyre), TNA (threose-nukleinsyre) og GNA (glyserol-nukleinsyre) har vært betraktet som mulige pre-RNAs.
23. M. Levy og S.L. Miller, 'The Stability of the RNA-bases', Proceedings of National Acadeym of Sciences 95 (1998), ss.7933-7938
25. W.K. Johnston et al, 'RNA-Catalysed RNA Polymerization ..', Science 292 (2001), s.1319
26. H.S. Zaher og P.J.Unrau, 'selection of an Improved RNA Polymeraze Ribozyme..' RNA 13 (200/), ss.1017-1026
27. J.W.Szostak et al, 'Synthesizing Life', Nature 409 (2001), ss.387-390
Oversatt og lagt til rette for .htm-format ved Asbjørn E. Lund